Mobility of group II introns
The major mobility event of group II introns is homing (or retrohoming), in which the intron inserts site-specifically into intronless sites. For organellar introns, this occurs during crosses between two strains, where one strain contains the intron and the other does not. For bacterial introns, homing occurs when homing site DNA is introduced into a cell expressing the intron.
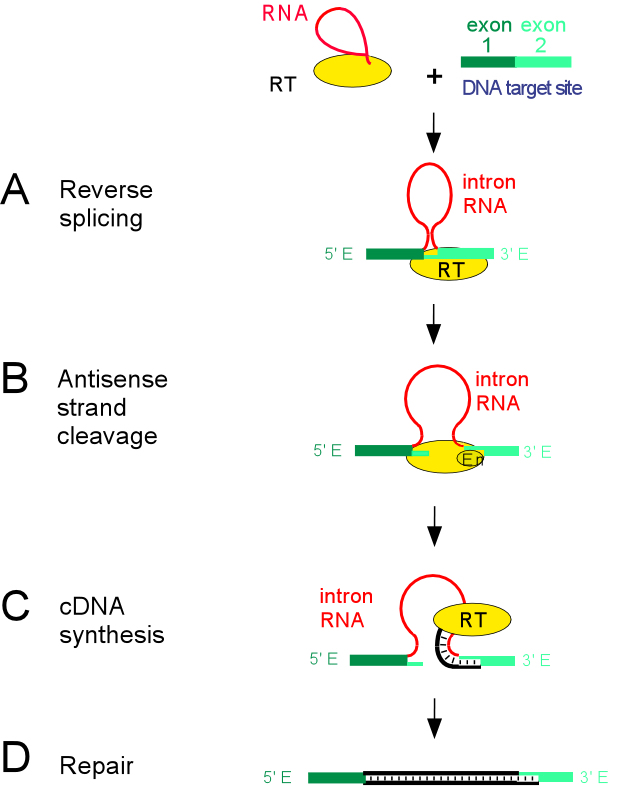
The mechanism of homing is termed target-primed reverse transcription, and is carried out by catalytic activities of both intron RNA and RT protein, with both subunits required for each reaction step.
The reaction begins by reverse splicing of the intron into the exon junction in double-stranded DNA. The reaction is RNA-catalyzed but facilitated by the RT protein, and is essentially the reverse of maturase-assisted splicing but occurs into DNA exons. Next, the En domain of the IEP cleaves the opposite strand 9 or 10 bp downstream, and the RT reverse transcribes the intron using the cleaved DNA as a primer. Recombination and repair activities complete the intron insertion process.
Many bacterial intron ORFs, including IIC ORFs, lack an En domain and as such rely on an endonuclease-independent pathway. In this case the primer for reverse transcription is provided by the replication fork during DNA replication.
At much lower frequencies, group II introns can insert into "unrelated", or ectopic, sites through retrotransposition. The mechanism of retrotransposition is believed to involve reverse splicing of the intron intro cryptic homing sites. Because intron insertion at illegitimate sites includes a step of reverse splicing into the target sequence, the intron should still be able to splice out of its new site.